Редагування РНК Зміст Загальнобіологічне значення | Історія відкриття | Типи редагування | Системи редагування | Мішені редагування | Редагування РНК і патології | Редагування РНК і еволюція | Джерела | Література | Посилання | Навігаційне менюMolecular diversity through RNA editing: a balancing actEvolutionary Origin of RNA Editing10.1021/bi300419rWidespread RNA and DNA Sequence Differences in the Human Transcriptome10.1126/science.1207018Very Few RNA and DNA Sequence Differences in the Human Transcriptome10.1371/journal.pone.00258422050-084X10.7554/eLife.051981360138510.1016/j.tplants.2016.07.005Rob Benne, Janny Van Den Burg, Just P.J. Brakenhoff, Paul Sloof, Jacques H. Van Boom, Marijke C. Tromp, Major transcript of the frameshifted coxll gene from trypanosome mitochondria contains four nucleotides that are not encoded in the DNA, Cell, Volume 46, Issue 6, 12 September 1986, Pages 819–826, Apolipoprotein B-48 is the product of a messenger RNA with an organ-specific in-frame stop codon10.1126/science.3659919RNA editing in brain controls a determinant of ion flow in glutamate-gated channels0092-867410.1016/0092-8674(91)90568-J0028-083610.1038/341660a00028-083610.1038/353178a00028-083610.1038/nature134000016-673110.1534/genetics.114.1614142045-232210.1038/s41598-017-04960-710.7717/peerj.152The Neurofibromatosis Type I Messenger RNA Undergoes Base-Modification RNA Editing10.1093/nar/24.3.478A novel translational repressor mRNA is edited extensively in livers containing tumors caused by the transgene expression of the apoB mRNA-editing enzyme10.1101/gad.11.3.3212494687010.1186/gb-2014-15-6-r790036-807510.1126/science.12127950737-403810.1093/molbev/msr1130265924710.1002/bies.200900045Молекулярна біологіяАполіпопротеїн В: особливості метаболізму в клітинах печінки телят при діареїМодифікація РНК для адаптаціїоригіналуМікроРНК теж піддаються редагуваннюоригіналуMolecular diversity through RNA editing: a balancing act0021-952510.1083/jcb.201511041Сайт присвячений Редагуванню РНКDARNED (DAtabase of RNa EDiting in humans, mice and flies). База даних з редагування РНК у людини, мишей і мухСайт з редагування A на IСайт з редагування C на Uр
Нуклеотидна послідовністьГенІнтронЕкзонГенетичний кодГеномФактори транскрипціїФактори термінації транскрипціїРНК-полімеразаIIIIIIПромоторВідкрита рамка зчитуванняОперонпре-мРНККепуванняСплайсингПоліаденілуванняРедагування РНКПсевдоуридинилюванняЕпігенетична регуляціяМетилювання ДНКCpG-острівціМетилювання гістонів[en]Ацетилювання та деацетилювання гістонів[en]Геномний імпринтингРегуляція транскрипції[en]Регуляторні послідовностіАктиваториЕнхансериРепресориРемоделювання хроматинуПост-транскрипційна регуляція[en]РНК-інтерференціямікроРНКАльтернативний сплайсингАльтернативне поліаденілуванняДеградація РНКПосттрансляційна регуляція[en]ФосфорилюванняСумоїлуванняГлікозилюванняУбіквітинуванняПротеоліз
Вибрані статтіРНКМолекулярна біологія
англ.нуклеотидівРНКтранскрипціїДНКсплайсингомкепуваннямполіаденілюваннямдеградацієютРНКтрансляціїмікроРНКевкаріотівбактерійархейтРНКнекодуючих РНКпротистрослинпам'яттюнавчаннямімунною відповіддю1986нідерландськаЛейденського університету«Cell»трипаносомцитохромоксидази IIурациловірамку зчитуваннятриплетного кодуМітохондріїкінетопластид1987Техасуаполіпопротеїн Bпечінціліпідівліпопротеїдівплазмі кровітонкого кишечникуцитозинглутамінстоп-кодонівГейдельберзького університетуглутаматні рецепториЕлектрофізіологічнінатріюкальціюAMPA-каїнатнихаргінінугіпермутагенезувірусівкоруангл.хлоропластахрису посівногокукурудзиметіонінтреонінкДНКсеквенуванняінозингуанозинЦНСдимерівдоменцинкуінозитол-гексафосфатуреброплависплайсингуAlu-елементівдальтонінтерферономнокауту генівМишіапоптозомкровотворенняепілептичнутрутовика лакованогоADATдріжджіваскоміцетівпсевдогеніA1CFB-субодиниці сукцинатдегідрогеназимоноцитахгіпоксіїNF1NAT1PPR-білківангл.покритонасіннихзеленими водоростямихлоропластівантоцеротовиднихпапоротейбомбезиннейропептидитрихоплаксапіридоксинслизовикаРНК-зв'язувальних білківвосьминогавалінпотенціалу діїтранспозонівінтронахекзонахполісомРНКазДрошаангл.Дайсерангл.антикодонісумчастихравликіввосьминогівлейшманійбактерійопосумаADATантикодоніоніхофордепресіїшизофреніїепілепсіїсуїцидальній поведінціпріоннихсиндрому Прадера-Віллібічному аміотрофічному склерозіхворобі Альцгеймерахворобі Гантінгтонасеротонінового рецептораонкологічних захворюванняхсупресорівпротоонкогенівастроцитомахгліомахгострого мієлоїдного лейкозураку молочної залозираку підшлункової залозилейкеміїВІЛвірус корувезикулярного стоматитуADATретротранспозиційPercolozoaгоризонтального переносу генівЮрій Чайковськийреплікацієюрекомбінацієюрепарацією ДНК

Взаємодія рибонуклеопротеїду (РНП), едитосоми (Е) і рибосоми під час редагування РНК у мітохондріях кінетопластид
Редагування РНК (англ. RNA editing, RNAe) — спрямована зміна окремих нуклеотидів у РНК після транскрипції[1]; клітинні процеси, що ведуть до модифікації РНК, яка не пов'язана з послідовністю матриці ДНК (чи РНК), з якої зчитана модифікована РНК[2]. Іншими словами, редагування РНК визначають як таку післятранскрипційну зміну РНК, яка не є сплайсингом, кепуванням, поліаденілюванням або деградацією. Результатом зміни нуклеотидів у РНК можуть бути зміни послідовності амінокислот і функції білка; поява чи зникнення сайтів сплайсингу мРНК; зміна послідовностей тРНК; відміна пригнічення трансляції за допомогою мікроРНК. Коливання рівня редагування РНК спостерігаються при деяких нервових захворюваннях, онкологічних та інфекційних патологіях.
Редагування РНК характерне майже виключно для евкаріотів, хоча в бактерій та архей виявлено декілька механізмів редагування тРНК. Різні групи евкаріотів мають різні механізми посттранскрипційної заміни нуклеотидів, що часто є унікальними для кожної групи[3].
Зміст
1 Загальнобіологічне значення
2 Історія відкриття
3 Типи редагування
4 Системи редагування
4.1 Редагування А на І
4.1.1 Аденозин-дезамінази РНК
4.1.2 Заміни А на І у грибів
4.2 Редагування Ц на У
4.2.1 Цитидиндезамінази
4.2.2 Заміни Ц на У в органелах вищих рослин
4.3 Редагування У на Ц
4.4 Редагування РНК у протист
5 Мішені редагування
5.1 Точкові заміни в екзонах
5.2 Редагування повторів
5.3 МікроРНК
5.4 Транспортні РНК
6 Редагування РНК і патології
6.1 Нервові захворювання
6.2 Онкологічні захворювання
6.3 Інфекційні захворювання
7 Редагування РНК і еволюція
7.1 Походження систем редагування РНК
7.2 Еволюційне значення редагування РНК
8 Джерела
9 Література
9.1 Україномовна
9.2 Англомовна
10 Посилання
Загальнобіологічне значення |
Редагування РНК є механізмом посттранскрипційних змін РНК. Цей процес разом з альтернативним сплайсингом збільшує різноманіття білків, які кодуються в ДНК, а також дозволяє розширити функціональну активність некодуючих РНК. Існують різні оцінки поширення редагування РНК у природі, тому і внесок процесу в регуляцію роботи клітини оцінюється спеціалістами неоднозначно. Одразу після відкриття редагування сприймалося як рідкісне явище, характерне для протист і рослин. Натомість 2010 року американська дослідниця Мінг'яо Лі повідомила, що у ссавців редагування зазнають більше як 95% транскриптів[4][5]. Надалі така оцінка піддавалася критиці молекулярних біологів як перебільшення[6]. До початку 2015 року у людини було надійно встановлено 115 сайтів редагування у мРНК за допомогою аденозин-дезаміназ, які призводили до заміни амінокислоти у білку, а також 53 сайти в мишей, тоді як дрозофіли мають 645 сайтів, а у нервовій системі кальмарів шляхом редагування РНК змінюються амінокислоти 60% білків.[7] Однак кількість виявлених випадків редагування РНК постійно зростає, особливо у регуляторних некодуючих ділянках РНК, що свідчить про непересічну роль механізму. Посилене редагування РНК у мозку та імунній системі ссавців вказує на те, що ця система регуляції роботи клітини забезпечує тонке налагодження процесів, пов'язаних із пам'яттю, навчанням, імунною відповіддю[8].
У рослин та грибів редагування РНК має адаптивне значення і призводить до виправлення помилок, які накопичуються в геномній ДНК ядра та органел.[9]
Історія відкриття |

Трипаносоми - перші організми, в яких виявлено редагування РНК
1986 року нідерландська група біохіміків під керівництвом професора Жака ван Бома з Лейденського університету опублікувала в журналі «Cell» дослідження мітохондріальних РНК одного з видів трипаносом[10][11]. Дослідники повідомили, що білок цитохромоксидази II відрізняється від передбачуваного продукту свого гену, починаючи зі 170-го амінокислотного залишку. Оскільки цей ген є висококонсервативним між різними видами трипаносом, вчені припустили, що він є функціонально важливим. Виявилось, що мРНК, яка зчитується з цього гену, має 4 додатково вставлені урацилові нуклеотиди, що й зсувало рамку зчитування триплетного коду. Нідерландські дослідники постулювали, що ці нуклеотиди вставляються після або під час транскрипції і є наслідком невідомого процесу, який вони назвали «редагуванням РНК».
Мітохондрії кінетопластид є унікальними органелами, тому дослідники були дуже обережними щодо екстраполяції свого відкриття на інші організми. Надалі виявилось, що більшість мітохондріальних мРНК трипаносом зазнають редагування зі вставлянням чи вилученням урацилів.
1987 року подібне явище було виявлено у людини. Група американських вчених із Техасу досліджувала білок аполіпопротеїн B, що існує у двох формах: ApoB-100 й ApoB-48[12]. Перша ізоформа за амінокислотною послідовністю збігалася з теоретично передбаченою з нуклеотидів гену ApoB, тоді як інша, менша за розміром, хоча й мала антигенні властивості N-кінцевої ділянки першої, проте значно відрізнялася за функцією. Більша форма ApoB-100 синтезується в печінці й відповідає за обмін ліпідів, перебуваючи у складі ліпопротеїдів великої і низької щільності в плазмі крові. Менша форма ApoB-48 секретується клітинами тонкого кишечнику. Аналіз мРНК у печінці й кишечнику виявив наявність повних транскриптів гену ApoB, але в кишечнику цитозин у 6457 положенні було замінено на урацил. Така заміна в цьому місці перетворювала кодон ЦАА, що кодує амінокислоту глутамін, на триплет УАА, що є одним зі стоп-кодонів. Це призводило до припинення трансляції білку й утворення вкороченої молекули ApoB-48. Автори дослідження не знали про роботу нідерландських біохіміків на трипаносомах, тому висунули гіпотезу про відкриття унікального механізму зміни транскрипції. У подальшому було описано цілу систему дезамінування цитозинів у мРНК після транскрипції.
Наступним кроком, що склав цілісне уявлення про процес редагування РНК, стала опублікована 1991 року в журналі Cell наукова праця німецької групи нейробіологів із Гейдельберзького університету, які досліджували глутаматні рецептори[13]. Електрофізіологічні експерименти довели, що існує два функціональних типи глутаматних рецепторів у мозку ссавців: одні, які пропускають лише іони натрію, та інші, проникні як для іонів натрію, так і для іонів кальцію. Селективність AMPA- і каїнатних глутаматних рецепторів забезпечується залишком аргініну або глутаміну в локусі на другому трансмембранному сегменті пептидного ланцюга. Дослідники виявили наявність як аргінінового, так і глутамінового кодону в мРНК глутаматних рецепторів, тоді як їхні гени містять лише триплети глутаміну. Заміна призводила до появи гуанінового нуклеотиду замість аденінового, тобто А змінювалося на Г. Відмінність між геномною ДНК і мРНК було пояснено саме явищем редагування РНК, до якого німецькі науковці вперше віднесли й нуклеотидні вставки в мітохондріях трипаносом, і появу стоп-кодону в аполіпопротеїні B. Це відкриття дало поштовх для дослідження системи аденіндезаміназ і всього розмаїття механізмів редагування РНК.
Аденозиндезамінази було відкрито наприкінці 1980-х років під час вивчення гіпермутагенезу вірусів кору. Було доведено, що при персистуванні інфекції в мозку значна кількість вірусної РНК мала велику кількість замін У на Ц[14]. Пізніше дослідники звернули увагу, що такі заміни відповідають мутаціям у РНК-матриці А на Г, і зіставили цей факт із можливою заміною А на І[15] Надалі було доведено, що така реакція справді має місце, здійснюється вона аденозиндезаміназою, що є інтерферон-залежною й дезамінує аденозини у дволанцюговій РНК (англ. DRADA, ds-RNA adenosine desaminase)[16].
У рослин редагування РНК у мітохондріях було вперше визначено у 1989 році в мітохондріях[17], а 1991 року в хлоропластах. Ген рибосомного білку L2 хлоропластів рису посівного та кукурудзи замість старт-кодону АУГ чи ГУГ, які кодують амінокислоту метіонін, мав триплет АЦГ (кодує треонін). Таким чином, трансляція білкового продукту була теоретично неможливою, оскільки синтез будь-якого білка починається з метіоніна, однак в реальності білок був виявлений в рибосомах хлоропластів. Німецькі дослідники припустили, що послідовність нуклеотидів у мРНК цього гену відрізняється від послідовності кільцевої ДНК хлоропласту, та дійсно виявили явище заміни Ц на У, яке призводило до появи старт-кодону.[18]
Типи редагування |
До відкриття нових сайтів редагування призводить накопичення інформації про послідовності ДНК і кДНК живих організмів. Усе кращі методи секвенування ДНК дозволяють отримати послідовність нуклеотидів, що містить мінімум помилок, а порівняння сиквенсу геномної ДНК і мРНК дає змогу знайти відмінності між цими послідовностями. У передгеномну еру такі відмінності пояснювали методичними помилками, але нові підходи «глибокого» багаторазового читання геномів і транскриптомів зводять можливі помилки до мінімуму. Таким чином, дослідження редагування РНК відбувається спочатку за результатами редагування в РНК, а вже потім вивчаються механізми таких редагувань. Тому для деяких типів редагування молекулярних систем досі не описано.
За якістю змін у послідовності РНК розрізняють такі типи редагування:
- вставки (інсерції) нуклеотидів
- видалення (делеції) нуклеотидів
- заміни нуклеотидів, що не призводять до зсуву рамки зчитування та не впливають на кількість нуклеотидів у РНК.
Найбільш поширеними серед усіх груп евкаріотичних організмів є саме нуклеотидні заміни:
- заміна А на І (або на Г) — найчастіше відбувається шляхом дезамінування аденозину (А) з утворенням інозину (І), нуклеозиду, який містить мінорну азотисту основу гіпоксантин, що розпізнається клітинними системами (трансляції, реплікації, репарації тощо) як гуанозин (Г); реакція здійснюється ферментами аденозин-дезаміназами
- заміна Ц на У — найчастіше відбувається шляхом дезамінування цитидину (Ц) з утворенням уридину (У); реакція здійснюється ферментами цитидиндезаміназами в тварин та PPR-білками в рослин
- заміни Г на А, У на Ц та деякі інші описано в літературі, але механізми для них поки що залишаються невідомими[1].
Системи редагування |
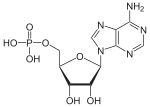
Аденозинмонофосфат

Інозинмонофосфат

Гуанозинмонофосфат
Редагування А на І |
Аденозин-дезамінази РНК |

Розмаїття, структура й гомологія генів родини ADAR. Дезаміназний домен — фіолетовий, РНК-зв'язуючий домен — жовтогарячий, Z-ДНК-зв'язуючий домен ADAR1 — зелений. Показано гени людини, кальмара, дрозофіли, нематоди, гідри, а також ADAT людини й дрозофіли
Аденозин-дезамінази РНК (ADAR) являють собою родину ферментів, які взаємодіють із дволанцюговою РНК, відщеплюють від аденозинового нуклеотиду NH2-групу та перетворюють його на інозин. За амінокислотною послідовністю й рентгенограмами білкових кристалів вони принципово відрізняються від інших аденозиндезаміназ (ADA), що відщеплюють аміногрупу від аденозинмонофосфату (АМФ). Деяку гомологію спостерігають між генами ADAR та цитозиндезаміназ, що може свідчити про їх походження від спільного гену-предка[19]. Утворений внаслідок дезамінування нуклеотид інозин розпізнається системами трансляції, РНК-інтерференції, зворотної транскрипції та іншими молекулярними машинами, побудованими на принципі комплементарності, як гуанозин.
До родини аденозин-дезаміназ у ссавців належать 3 гени: ADAR1, ADAR2, ADAR3. Білок ADAR3 присутній лише в головному мозку, тоді як ADAR1 і ADAR2 наявні в усіх тканинах, хоча більше їх також у ЦНС. Аденозин-дезамінази діють у формі димерів. Кожен із білків ADAR має у своєму складі РНК-зв'язуючий домен, який містить іон цинку. Також в активному центрі ферменту є молекула інозитол-гексафосфату (IP6).
Найбільшу кількість генів ADAR станом на 2018 рік мають реброплави, у геномі яких наявні 4 ізоформи аденозиндезаміназ. Ці ферменти є найбільш активними під час ембріогенезу.[20]
Аденозин-дезамінази взаємодіють із первинними транскриптами, найчастіше — до сплайсингу РНК. Мішенню є дволанцюгова РНК, зазвичай — спарені нуклеотиди у шпильках. Невідомо, чи існують послідовності, що є сприятливими до редагування, але є відомості щодо специфічної третинної й четвертинної укладки молекули РНК, яку використовують аденозин-дезамінази. Редагування відбувається в пре-мРНК, первинних транскриптах мікроРНК, у транскриптах повторів (наприклад, Alu-елементів). Редагування призводить до замін амінокислот у білку, що синтезується з мРНК, до створення або ліквідації сайтів для сплайсингу чи до зміни активності мікроРНК.
Вкорочений сплайс-варіант ферменту ADAR1 і білок ADAR2 містяться в ядрі та експресуються постійно. Білок ADAR1 із повною послідовністю (p150, масою 150 кілодальтон) рухається з ядра до цитоплазми й назад. Для білку ADAR3 людини не доведено дезаміназної активності[1].
Редагування А на І може відбуватися для всіх транскриптів (Q/R сайт глутаматних рецепторів — понад 99,9% транскриптів редагуються) або лише для частини з них, тоді як інші транскрипти залишаються нередагованими. Регуляція редагування здійснюється багатьма факторами: інтерфероном, транскрипційними кофакторами, збиранням субодиниць білка в димери тощо. ADAR2 саморегулюється через негативний зворотний зв'язок: надлишок білка починає редагувати пре-мРНК самого ADAR2, викликаючи появу додаткового сайту сплайсингу й утворення вкороченого нефункціонального ферменту.
Досліди з нокауту генів аденозин-дезаміназ довели, що вони є життєво важливими для ссавців. Миші з вимкненим ADAR1 гинули ще в ембріональний період розвитку через масову втрату нейронів апоптозом і порушення системи кровотворення. Миші, позбавлені ADAR2, помирали одразу після народження через епілептичну активність мозку. Проте заміна лише одного нуклеотиду А на Г у сайті гену GluR2, що кодує глутамін, надавала змогу нокаутним мишам жити без таких порушень[1].
Заміни А на І у грибів |
У 2014 році у ядерному геномі трутовика лакованого було виявлено численні сайти редагування РНК, зокрема з заміною А на І, вперше серед вищих грибів. При цьому жодного ферменту, гомологічного білкам ADAR тварин, у геномі цього трутовика не виявлено. Втім, знайдено 3 гени, споріднені з дезаміназами родини ADAT, близькі до даких у дріжджів. Більшість замін спостерігалося в кодуючих послідовність білка ділянках ДНК.[21]
Детальне вивчення редагування А на І у аскоміцетів роду Fusarium виявило їх роль у статевому розмноженні та спороутворенні. Зокрема редагування одного з нуклеотидів у псевдогені AMD прибирає стоп-кодон і відновлює синтез білка, який відповідає за дозрівання та відокремлення спор.[22]
Редагування Ц на У |
Цитидиндезамінази |

Реакція дезамінування цитозину
До родини цитидиндезаміназ належать білки AID/APOBEC, що здатні дезамінувати цитидин як у складі РНК, так і в ДНК. Головну роль у редагуванні РНК відіграє фермент APOBEC1. Якщо аденозиндезамінази взаємодіють із мРНК до чи під час сплайсингу, APOBEC1 редагує вже зрілі мРНК в ядрі. Для редагування необхідний білковий комплекс, до якого окрім APOBEC1 входить білок A1CF (apobec-1 complementation factor), допоміжний фактор APOBEC1 і ще деякі білки. Цей комплекс розпізнає ділянку РНК у 30-40 нуклеотидів, що утворює шпильку, причому цільовий цитозин міститься в «голівці» (неспареній ділянці) цієї шпильки. Білок A1CF виконує адапторну функцію, забезпечуючи контакт дезамінази з РНК. Комплекс розпізнає послідовність UUUN(A/U)U, яка має міститися далі за сайтом редагування та перетинається з якірною послідовністю, до якої приєднується молекула A1CF.
Від ступеня збігу цієї ділянки різних РНК з послідовністю мРНК ApoB залежить ефективність редагування конкретного цитидину.
На відміну від аденозиндезаміназ, у ссавців станом на 2013 рік було відомо лише чотири сайти редагування. Окрім транскрипту ApoB, редагується мРНК B-субодиниці сукцинатдегідрогенази мітохондрій у моноцитах людини, причому редагування активізується при гіпоксії[23], транскрипт гену фактору нейрофіброматозу NF1,[24] і мРНК репресору трансляції NAT1[25]. У всіх цих випадках редагування зі звичайного кодону, який відповідає амінокислоті, утворюються стоп-кодони, що призводить до синтезу вкороченої форми білка, яка може бути функціональною (як у ApoB) чи нефункціональною (при редагуванні NAT-1). Проте дослідження 2014 року виявило ще більше 50 подібних сайтів у кишечнику і печінці мишей. [26]
Заміни Ц на У в органелах вищих рослин |
У мітохондріях і хлоропластах судинних рослин поширене редагування мРНК і тРНК. Найчастіше заміни Ц на У відбуваються в кодонах мРНК, причому характерне редагування першого чи другого нуклеотиду триплету, яке веде до заміни амінокислоти. У багатьох мітохондріях вищих рослин наявні до 300–500 сайтів для редагування РНК. Виявлено групу PPR-білків (англ. pentatricopeptide repeat), що відповідають за розпізнавання сайтів редагування. Родина цих білків у покритонасінних є однією з найбільших і містить від 400 до 600 білків, проти 10-20 у всіх інших евкаріотів, включно із зеленими водоростями[3].
Основною функцією такого редагування вважається не збільшення білкового різноманіття, а виправлення мутацій Т→Ц у геномі органел. Спостерігається тенденція до зниження числа сайтів редагування в органелах вищих покритонасінних порівняно з голонасінними та папоротеподібними. Це пояснюють зворотними мутаціями Ц→Т в ДНК мітохондрій та пластид.[9]
Редагування У на Ц |

Механізм вставки додаткових урацилів до мРНК при редагуванні РНК у трипаносом. гРНК - гідова РНК
Процес, характерний здебільшого для мітохондрій і хлоропластів антоцеротовидних мохів та папоротей. Крім того, задокументовано подібні випадки у РНК-транскриптів, що кодують бомбезин-подібні нейропептиди в амфібій, у генах WT1 ссавців, а також у мітохондріальній мРНК трихоплакса. Механізми такої заміни невідомі, але припускають, що вони можуть бути подібними до реакції, яка каталізується ферментом ссавців ЦТФ-синтазою, піридоксин-залежним ензимом, що відіграє ключову роль у метаболізмі нуклеотидів, переносячи аміногрупу з глутаміну до УТФ для синтезу ЦТФ.
Редагування РНК у протист |
Вставки і видалення нуклеотидів у кінетопластид відбуваються майже в усіх мітохондріальних транскриптах. Видаляються і вставляються уридинові залишки з пре-мРНК, що регулюється короткими гідовими РНК, які є комплементарними до вже відредагованої мРНК. Особливий багатобілковий комплекс (едитосома) розрізає пре-мРНК і вставляє або видаляє уридини залежно від комплементарної послідовності гідової РНК.
Інший тип вставок нуклеотидів спостерігають у мітохондріях слизовика Physarum polycephalum. У цьому випадку редагування призводить до вставки додаткових цитозинів під час транскрипції.
Мішені редагування |
Основна частина редагування РНК у тварин відбувається шляхом заміни А на І за участі аденозин-дезаміназ. Відомо лише 4 сайти в РНК для цитозин-дезаміназ. Редагування у тРНК досить різноманітне в різних групах організмів, але там теж переважає дезамінування аденіну.
Точкові заміни в екзонах |
Станом на початок 2015 року у людини було надійно встановлено 115 сайтів редагування у мРНК за допомогою аденозин-дезаміназ, які призводили до заміни амінокислоти у білку, а також 53 сайти в мишей, тоді як дрозофіли мають 645 сайтів, а у нервовій системі кальмарів шляхом редагування РНК змінюються амінокислоти 60% білків.[7]. Найбільший рівень редагування спостерігається у мРНК іонних каналів і рецепторів мозку, рецепторів імунної системи, білків цитоскелету, РНК-зв'язувальних білків[8]. Деякі дослідники вважають, що найбільш часто редагуються ті мРНК, які є довгими, а значить мають складнішу третинну структуру і більше шпильок.[7]
Цікавим прикладом редагування є заміни А на І у мРНК калієвого каналу Kv1 восьминога. У восьми досліджених видів із тропічних та полярних океанічних вод рівень редагування цієї мРНК корелює зі зниженням температури. Замінена під час редагування амінокислота валін дає змогу каналу швидше інактивуватися, тому для досягнення нового потенціалу дії потрібно менше часу, що вигідно в умовах зниженої температури середовища[27].
| Ген | Амінокислотна заміна | Функція білка | Тканина | Результат редагування |
|---|---|---|---|---|
GluR2 (Gria2, GluR-B, AMPA2) | Q/R R/G | Глутаматний AMPA-рецептор з іонним каналом | мозок | Непроникність каналу для кальцію |
| GluR3 | R/G | Глутаматний AMPA-рецептор з іонним каналом | мозок | Прискорене відновлення після десенсетизації |
| GluR4 | R/G | Глутаматний AMPA-рецептор з іонним каналом | мозок | Прискорене відновлення після десенсетизації |
GluR5 (Grik1) | Q/R | Глутаматний каїнатний рецептор з іонним каналом | мозок | Непроникність каналу для кальцію |
GluR6 (Grik2) | Q/R I/V Y/C | Глутаматний каїнатний рецептор з іонним каналом | мозок | Зміна проникності для іонів |
| KCNA1 | I/V | Потенціал-залежний калієвий канал | мозок | Уповільнена інактивація каналу |
| Gabra3 | K/E | Рецептор ГАМК з іонним каналом | мозок, кишечник | Зміна спорідненості до ГАМК |
| 5HT2cR | I/V I/M N/S N/D N/G I/V | Серотоніновий рецептор | мозок | Змінене зв'язування із серотоніном, змінений транспорт рецептора |
| CyFIP2 | I/M | Регулятор полімеризації актину | всюди | ? |
| FLNA | Q/R | Філамін А, взаємодіє з актином у розгалуженнях, місцях прикріплення рецепторів, при передачі сигналів | всюди, редагування активне у шлунково-кишковому тракті | можливий сплайсинг |
| FLNB | Q/R | Філамін B, взаємодіє з актином у розгалуженнях, місцях прикріплення рецепторів, при передачі сигналів | всюди, редагування активне у шлунково-кишковому тракті | ? |
| BLCAP | K/R Q/R Y/C | Зменшена активність при метастатичному раку сечового міхура та при інших пухлинах | сечовий міхур | ? |
| IGFBP7 | K/R R/G | Інсуліновий рецептор, регуляція росту й міграції клітин гліоми | мозок, глія | змінює зв'язування з гепарином |
| C1QL1 | T/A Q/R Q/R | Елемент системи комплементу | лімфоцити, макрофаги, клітини Пуркіньє мозочка | зміни олігомеризації білка |
| HMCN1 | K/E | Білок позаклітинного матриксу | око | ? |
| RSU1 | M/V | Білок клітинної адгезії, міграції | ? | |
| KCNMA1 | S/G | Кальційзалежний калієвий канал великої провідності | ? | |
| KIF20B | K/R | Кінезиновий білок, транспорт по мікротрубках, цитокінез | всюди | ? |
| ARFGAP2 | Q/R | Транспортний білок COPI (coat protein complex 1) | ? | |
| CD6 | S/G | Поверхневий рецептор активації Т-лімфоцитів | Т-лімфоцити | ? |
| GANAB | Q/R | Глюкозидаза, відрізає N-зв'язані глікани від білків | ? | |
| OS9 | E/G | Глікопротеїн системи деградації, пов'язаної з ретикулумом | ? | |
| NEIL1 | K/R | Специфічна до окиснювального пошкодження ендонуклеаза, ДНК-гликозилаза | всюди | зміна специфічності ферменту |
| MEX3B | Q/R | Локалізований у P-тільцях білок, РНК-зв'язуючий, регуляція мРНК | ? | |
STRN4 (стріатин, зінедин) | R/G | Білок цитоскелету, кальмодулін-зв'язуючий білок, передача сигналу в нейронах | нейрони | ? |
BIN1 (амфіфізин II) | K/R | Пухлинний супресор, кардіоміопатія | мозок | ? |
| SS18L1 | S/G | Кальційзалежний активатор транскрипції, необхідний для нормального росту й галуження дендритів у нейронах кори | нейрони кори головного мозку | ? |
| CADPS | E/G | Кальційзалежний активатор секреції й екзоцитозу щільних везикул | ? | |
ATXN7 (атаксин 7) | K/R | Субодиниця ацетил-трансферази гістонів | мозок | ? |
| TTLL3 | K/R | Представник родини подібних до тубулінових тирозин-лігаз білків, необхідний для роботи війок | кишечник | ? |
CCNI (циклін 1) | R/G | Антиапоптичний фактор, регулятор клітинного циклу | клітини одразу після мітозу | ? |
| PTPRN2 | E/G | Тирозинова фосфатаза, трансмембранний білок у щільних секреторних везикулах у нейроендокринних клітинах, автоантиген при діабеті І типу, бере участь у регуляції поведінки, тривалості життя, секреції нейромедіаторів | нейрони | ? |
| AZIN1 | S/G | Інгібітор орнітин-декарбоксилази | підвищене редагування при раку печінки | |
PTK2 (тирозинова протеїн-кіназа 2) | T/A | Пов'язана з цитоскелетом протеїн-кіназа | ? | |
| FBXL6 | Stop/W | Убіквітин-лігаза білка Tel (ETV6) | ? | |
| CRB2 | T/A | Трансмембранний білок, бере участь у міжклітинних контактах | ? | |
TRO (трофінін) | S/G | Трансмембранний білок, білок клітинної адгезії, імплантація ембріону у ссавців | ? | |
| ARL6IP4 | K/R | Гомолог фактору сплайсингу | ? | |
| Elavl2 | I/V N/D | Нейрон-специфічний РНК-зв'язувальний білок, диференціація нейронів і синаптична пластичність | нейрони | ? |
Редагування повторів |
У ссавців редагування шляхом дезамінування аденозину найчастіше трапляється в повторах, особливо в таких, що походять із транспозонів. У геномі приматів багато таких повторів мають Alu-елементи, що через мале внутрішнє різноманіття й високу комплементарність один до одного створюють подвійні ланцюги РНК у транскриптах. Такі дволанцюгові ділянки РНК є ідеальним місцем для зв'язування ферментів ADAR та вони часто зазнають множинного редагування А на І.
Більшість Alu-елементів, як й інших повторів, містяться в інтронах генів або в нетрансльованих екзонах, тому їх редагування не змінює послідовність амінокислот білка. Попри це, редагування може впливати на появу або зникнення сайтів сплайсингу. Зокрема, редагування А на І може створювати донорний (АТ на ІТ) або акцепторний (АА на АІ) сайт сплайсингу, змінювати ефективність сплайсингу за рахунок редагування енхансерної або інгібіторної послідовності на РНК, руйнувати акцепторний сайт сплайсингу (АГ на ІГ). Більшість же Alu-елементів зазнають масового редагування, що призводить до появи великого вмісту інозину в їх послідовностях. Є відомості, що частина таких багатих на інозин РНК руйнується в ядрі за допомогою спеціального інозин-чутливого білкового комплексу, хоча інші подібні РНК знайдено в цитоплазмі навіть у складі полісом.
МікроРНК |
МікроРНК походять із пре-мікроРНК, що являє собою дволанцюгову РНК-шпильку. Тому вони також є звичайним субстратом для роботи аденозин-дезаміназ. Редагування може запобігти дозріванню мікроРНК, якщо відредаговано сайти для РНКаз Дроша (англ. Drosha) і Дайсер (англ. Dicer). Заміни А на І у посадковій послідовності мікроРНК, яка зв'язується із сайтом на цільовій мРНК і викликає блокування її трансляції, можуть призводити до зміни репертуару матричних РНК, на які діє конкретна нередагована мікроРНК. Це характерно, наприклад, до мікроРНК-376 у людини. Редагування також може змінювати самі сайти для мікроРНК на мРНК, що знов-таки змінює подальшу РНК-інтерференцію.
Транспортні РНК |
Транспортні РНК мають цілий набір змінених нуклеотидів із мінорними азотистими основами у складі (таких нуклеотидів у тРНК зустрічається 79 типів). Редагування тРНК замінює нуклеотиди в антикодоні на канонічні плюс інозин. Такі заміни доведено для Acanthamoeba, Spizellomyces, слизовиків, сумчастих, наземних рослин, равликів, восьминогів, лейшманій, бактерій. Наприклад, в опосума редагування антикодону «аспартатної» тРНК у мітохондріях призводить до її здатності приєднувати гліцин[28]. Редагування створює також необхідні елементи для утворення вторинної, третинної і четвертинної структури тРНК, включаючи петлі й спарені нуклеотиди шпильок. Це здійснюється за рахунок як вставок/видалень, так і шляхом заміни нуклеотидів.
Для дезамінування аденозину в тРНК існує окрема підродина ферментів ADAT, аденозиндезаміназ транспортних РНК. Основні заміни відбуваються в антикодоні та акцепторній послідовності.
Найбільш щільне редагування тРНК відбувається в мітохондріях оніхофор. У них редагується до 50% нуклеотидів[29].
Редагування РНК і патології |

Зростання кількості тематичних наукових статей за даними біомедичної бази даних PubMed у 1986–2012 роках. Пошук за ключовими словами «RNA+editing» (РНК+редагування)
Нервові захворювання |
Активність редагування РНК у нервовій системі підвищена й відіграє важливу роль у її функціонуванні, тому ризики появи патологічних дефектів редагування тут більші за інші тканини. Змінене редагування РНК виявлено при деяких випадках депресії, шизофренії, епілепсії, суїцидальній поведінці, пріонних нейродегенеративних захворюваннях, аутосомній домінантній катапсії І типу, синдрому Прадера-Віллі, бічному аміотрофічному склерозі, хворобі Альцгеймера, хворобі Гантінгтона[1].
Під час бічного аміотрофічного склерозу спостерігається знижене редагування Q/R-сайту глутаматних рецепторів, що ймовірно призводить до підвищеного входу кальцію у нейрони та їх загибелі. Помилкове редагування серотонінового рецептора доведено для цілого переліку захворювань нервової системи. Таким чином, ферменти ADAR є потенційними мішенями для лікування багатьох психічних і неврологічних порушень.
Онкологічні захворювання |
Роль редагування РНК під час онкологічних захворюваннях є нез'ясованою. Вважається, що помилкова зміна нуклеотидів у мРНК і мікроРНК може призводити до інактивації супресорів пухлин або до активації протоонкогенів.
Порушення редагування мРНК спостерігаються під час деяких типів раку. Загальний рівень редагування РНК зменшується, особливо при астроцитомах та гліомах. Під час гострого мієлоїдного лейкозу виявлено неправильне редагування мРНК, що викликає неканонічний сплайсинг та утворення нефункціонального білку тирозинової кінази PTPN6. Під час досліджень раку молочної залози виявлено групу нових сайтів редагування мРНК, хоча ці сайти редагувалися і в нормальних клітинах[1].
Також виявлено вплив замін А на І у мікроРНК, наприклад, зменшення редагування мікроРНК-376 спостерігалося під час раку підшлункової залози, а мікроРНК-142 — під час лейкемії[1]. Допоки незрозуміло, чи зміни редагування передують злоякісній трансформації клітин, чи навпаки — викликаються нею.
Інфекційні захворювання |
Білки ADAR1 і APOBEC є активними регуляторами антивірусного імунітету. Вони активуються інтерфероном та здатні редагувати як РНК, так і ДНК вірусів, блокуючи їх розмноження ще з цитоплазми. Вірогідно, аденозин-дезамінази є також супресорами антивірусної відповіді, тому деякі віруси здатні використовувати ADAR1 для блокування апоптозу й імунної відповіді, зокрема ВІЛ, вірус кору, вірус везикулярного стоматиту[1].
Редагування РНК і еволюція |

Наявність генів ADAR у тварин. Червоним позначено ADAR1, синім — ADAR2. Видно, що обидва гени з'явилися ще до розділення багатоклітинних тварин, але комахи, ракоподібні й нематоди втратили ADAR1
Походження систем редагування РНК |
Згідно із сучасними уявленнями, системи редагування РНК мають поліфілетичне походження.[3].
Білки родин ADAR і AID/APOBEC, вірогідно, походять від предкового білку з родини ADAT, який виконував функцію редагування тРНК. Інша гіпотеза передбачає їхнє походження від бактеріальних дезаміназ токсинів.
Білки PPR вищих рослин, імовірно, походять від спільних для всіх евкаріотів предкових генів, що розмножилися у геномі за рахунок множинних ретротранспозицій. 2010 року подібний до рослинного тип редагування у мітохондріях виявлено у протисти Naegleria gruberi з типу Percolozoa. Залишається невідомим, чи є це прикладом горизонтального переносу генів, чи є спільність походження цієї системи.
Вставки уридинів, що спостерігаються у мітохондріях кінетопластид, властиві лише представникам цього класу, хоча схожі події у мітохондріальному транскриптомі описано у представника спорідненого класу диплонеміди Diplonema papillatum.
Еволюційне значення редагування РНК |
Різноманітні системи редагування РНК виникали багаторазово в ході еволюції та існують у більшості відомих великих таксонів. Змінюючи інформацію, що надходить із ДНК, вони дозволяють на базі одного генотипу існувати великому розмаїттю функціональних білків. Редагування А на І у ссавців часто відбувається в консервативних сайтах білку, де повна заміна амінокислоти має летальний ефект. Проте часткове чи тканиноспецифічне редагування таких сайтів дозволяє існувати організмам, що мають у своєму складі такі видозмінені білки. При певних змінах умов навколишнього середовища отримані в такий спосіб варіанти ферментів можуть виявитися корисними[30].
Російський історик науки Юрій Чайковський вважає процес редагування РНК та його виникнення центральним для розуміння біологічної еволюції нарівні з реплікацією, рекомбінацією та репарацією ДНК[31].
Особливістю ссавців є вища експресія ферментів редагування РНК у найбільш прогресивних системах органів: нервовій та імунній. Система аденозин-дезаміназ є дуже чутливою до зовнішніх впливів, тому може відігравати важливу роль в еволюції цих систем. Найбільший рівень редагування А на І у РНК серед хребетних характерний для людини. Alu-елементи людини й приматів мають дуже високий ступінь внутрішньої подібності порівняно з аналогічними повторами інших ссавців, тому їх транскрипти частіше утворюють дволанцюгові структури, придатні до редагування РНК. Разом із високою аденозин-дезаміназною активністю в мозку цей факт дає деяким дослідникам можливість припускати особливу роль редагування в еволюції людини[30].
Джерела |
↑ абвгдежи Farajollahi, Sanaz and Maas, Stefan (2010). Molecular diversity through RNA editing: a balancing act. Trends in Genetics (Elsevier) 26 (5): 221–230.
↑ Gott JM, Emeson RB. Functions and mechanisms of RNA editing. Annu Rev Genet 2000;34:499-531. PubMed:11092837
↑ абв Gray, Michael W. (2012). Evolutionary Origin of RNA Editing. Biochemistry 51 (26): 5235–5242. doi:10.1021/bi300419r.
↑ http://www.nature.com/news/2010/101105/full/news.2010.586.html
↑ Mingyao Li, Isabel X. Wang, Yun Li, Alan Bruzel, Allison L. Richards, Jonathan M. Toung, and Vivian G. Cheung (2011). Widespread RNA and DNA Sequence Differences in the Human Transcriptome. Science 333 (6038): 53–58. doi:10.1126/science.1207018.
↑ Schrider DR, Gout J-F, Hahn MW (2011). Very Few RNA and DNA Sequence Differences in the Human Transcriptome. PLoS ONE 6 (10). doi:10.1371/journal.pone.0025842.
↑ абв Alon, Shahar; Garrett, Sandra C; Levanon, Erez Y; Olson, Sara; Graveley, Brenton R; Rosenthal, Joshua J C; Eisenberg, Eli (2015). The majority of transcripts in the squid nervous system are extensively recoded by A-to-I RNA editing. eLife 4. ISSN 2050-084X. doi:10.7554/eLife.05198.
↑ аб Pullirsch D, Jantsch MF (2010). Proteome diversification by adenosine to inosine RNA-editing. RNA Biol 7: 1–8.
↑ аб Sun, Tao; Bentolila, Stephane; Hanson, Maureen R. (2016). The Unexpected Diversity of Plant Organelle RNA Editosomes. Trends in Plant Science 21 (11): 962–973. ISSN 13601385. doi:10.1016/j.tplants.2016.07.005.
↑ http://www.research.leiden.edu/famous/boom.html
↑ Rob Benne, Janny Van Den Burg, Just P.J. Brakenhoff, Paul Sloof, Jacques H. Van Boom, Marijke C. Tromp, Major transcript of the frameshifted coxll gene from trypanosome mitochondria contains four nucleotides that are not encoded in the DNA, Cell, Volume 46, Issue 6, 12 September 1986, Pages 819–826, ISSN [https://www.worldcat.org/search?fq=x0:jrnl&q=n2:0092-8674 0092-8674, http://dx.doi.org/10.1016/0092-8674(86)90063-2].
↑ Chen, SH and Habib, G and Yang, CY and Gu, ZW and Lee, BR and Weng, SA and Silberman and Cai, SJ and Deslypere, JP and Rosseneu, M and et, al. (1987). Apolipoprotein B-48 is the product of a messenger RNA with an organ-specific in-frame stop codon. Science 238 (4825): 363–366. doi:10.1126/science.3659919.
↑ Bernd Sommer and Martin Köhler and Rolf Sprengel and Peter H. Seeburg (1991). RNA editing in brain controls a determinant of ion flow in glutamate-gated channels. Cell 67 (1): 11–19. ISSN 0092-8674. doi:10.1016/0092-8674(91)90568-J.
↑ Cattaneo R, Schmid A, Eschle D, Baczko K, ter Meulen V, Billeter MA. Biased hypermutation and other genetic changes in defective measles viruses in human brain infections. Cell. 1988 Oct 21;55(2):255-265.
↑ Bass BL, Weintraub H, Cattaneo R, Billeter MA (1989) Biased hypermutation of viral RNA genomes could be due to unwinding/modification of double-stranded RNA. Cell 56: 331
↑ Weier, H. U. G., George, C. X., Greulich, K. M., & Samuel, C. E. (1995). The interferon-inducible, double-stranded RNA-specific adenosine deaminase gene (DSRAD) maps to human chromosome 1q21. 1-21.2. Genomics, 30(2), 372–375.
↑ Gualberto, José M.; Lamattina, Lorenzo; Bonnard, Géraldine; Weil, Jacques-Henry; Grienenberger, Jean-Michel (1989). RNA editing in wheat mitochondria results in the conservation of protein sequences. Nature 341 (6243): 660–662. ISSN 0028-0836. doi:10.1038/341660a0.
↑ Hoch, Brigrtte; Maier, Rainer M.; Appel, Kurt; Igloi, Gabor L.; Kössel, Hans (1991). Editing of a chloroplast mRNA by creation of an initiation codon. Nature 353 (6340): 178–180. ISSN 0028-0836. doi:10.1038/353178a0.
↑ Willemijn M. Gommans, Dylan E. Dupuis, Jill E. McCane, Nicholas E. Tatalias, Stefan Maas (2008). Diversifying Exon Code through A-to-I RNA Editing. У Smith, H. DNA RNA Editing. Wiley & Sons, Inc. с. 3–30.
↑ Moroz, Leonid L.; Kocot, Kevin M.; Citarella, Mathew R.; Dosung, Sohn; Norekian, Tigran P.; Povolotskaya, Inna S.; Grigorenko, Anastasia P.; Dailey, Christopher; Berezikov, Eugene; Buckley, Katherine M.; Ptitsyn, Andrey; Reshetov, Denis; Mukherjee, Krishanu; Moroz, Tatiana P.; Bobkova, Yelena; Yu, Fahong; Kapitonov, Vladimir V.; Jurka, Jerzy; Bobkov, Yuri V.; Swore, Joshua J.; Girardo, David O.; Fodor, Alexander; Gusev, Fedor; Sanford, Rachel; Bruders, Rebecca; Kittler, Ellen; Mills, Claudia E.; Rast, Jonathan P.; Derelle, Romain; Solovyev, Victor V.; Kondrashov, Fyodor A.; Swalla, Billie J.; Sweedler, Jonathan V.; Rogaev, Evgeny I.; Halanych, Kenneth M.; Kohn, Andrea B. (2014). The ctenophore genome and the evolutionary origins of neural systems. Nature 510 (7503): 109–114. ISSN 0028-0836. doi:10.1038/nature13400.
↑ Zhu, Yingjie; Luo, Hongmei; Zhang, Xin; Song, Jingyuan; Sun, Chao; Ji, Aijia; Xu, Jiang; Chen, Shilin (2014). Abundant and Selective RNA-Editing Events in the Medicinal Mushroom Ganoderma lucidum. Genetics 196 (4): 1047–1057. ISSN 0016-6731. doi:10.1534/genetics.114.161414.
↑ Cao, Shulin; He, Yi; Hao, Chaofeng; Xu, Yan; Zhang, Hongchang; Wang, Chenfang; Liu, Huiquan; Xu, Jin-Rong (2017). RNA editing of the AMD1 gene is important for ascus maturation and ascospore discharge in Fusarium graminearum. Scientific Reports 7 (1). ISSN 2045-2322. doi:10.1038/s41598-017-04960-7.
↑ Baysal, Bora E. and De Jong, Kitty and Liu, Biao and Wang, Jianmin and Patnaik, Santosh K. and Wallace, Paul K. and Taggart, Robert T. (9 2013). Hypoxia-inducible C-to-U coding RNA editing downregulates SDHB in monocytes,. doi:10.7717/peerj.152.
↑ Gary R. Skuse,; Amedeo J. Cappione, Mark Sowden, Linda J. Metheny, and Harold C. Smith (1996). The Neurofibromatosis Type I Messenger RNA Undergoes Base-Modification RNA Editing. Nucl. Acids Res. 24 (3): 478–486. doi:10.1093/nar/24.3.478.
↑ S Yamanaka; K S Poksay, K S Arnold, and T L Innerarity (February 1, 1997). A novel translational repressor mRNA is edited extensively in livers containing tumors caused by the transgene expression of the apoB mRNA-editing enzyme. Genes Dev. 11: 321–333. doi:10.1101/gad.11.3.321.
↑ Valerie Blanc, Eddie Park, Sabine Schaefer, Melanie Miller, Yiing Lin, Susan Kennedy, Anja M. Billing, Hisham Ben Hamidane, Johannes Graumann, Ali Mortazavi, Joseph H. Nadeau & Nicholas O. Davidson (2014). Genome-wide identification and functional analysis of Apobec-1-mediated C-to-U RNA editing in mouse small intestine and liver. Genome biology 15 (6): R79. PMID 24946870. doi:10.1186/gb-2014-15-6-r79.
↑ Garrett S.; Rosenthal J. J. C. (2012). RNA Editing Underlies Temperature Adaptation in K+ Channels from Polar Octopuses. Science 335 (6070): 848–851. ISSN 0036-8075. doi:10.1126/science.1212795.
↑ Börner, GV and Mörl, M and Janke, Axel and Pääbo, S (1996). RNA editing changes the identity of a mitochondrial tRNA in marsupials.. The EMBO journal (Nature Publishing Group) 15 (21): 5949.
↑ Segovia R.; Pett, W.; Trewick, S.; Lavrov, D. V. (2011). Extensive and Evolutionarily Persistent Mitochondrial tRNA Editing in Velvet Worms (Phylum Onychophora). Molecular Biology and Evolution 28 (10): 2873–2881. ISSN 0737-4038. doi:10.1093/molbev/msr113.
↑ аб Gommans Willemijn M., Mullen Sean P., Maas Stefan (2009). RNA editing: a driving force for adaptive evolution?. BioEssays 31 (10): 1137–1145. ISSN 02659247. doi:10.1002/bies.200900045.
↑ Ю. В. Чайковский (2003). Эволюция. М: Товарищество КМК.
Література |
Україномовна |
А. В. Сиволоб (2008). Молекулярна біологія. К: Видавничо-поліграфічний центр «Київський університет». с. 194.
Л. Калачнюк, О. Савка, Д. Мельничук, Г. Калачнюк (2004). Аполіпопротеїн В: особливості метаболізму в клітинах печінки телят при діареї. Вісник Львівського університету. Серія біологічна (українська) 38: 57–66.
Мітч Леслі (2012-01-10). Модифікація РНК для адаптації. Український науковий клуб. Архів оригіналу за 2014-01-10.
Сергєєв, Олександр. МікроРНК теж піддаються редагуванню. Наука. Технология. Архів оригіналу за 2014-01-10.
Англомовна |
Farajollahi, Sanaz and Maas, Stefan (2010). Molecular diversity through RNA editing: a balancing act. Trends in Genetics (Elsevier) 26 (5): 221–230.
Licht, Konstantin; Jantsch, Michael F. (2016). Rapid and dynamic transcriptome regulation by RNA editing and RNA modifications. The Journal of Cell Biology 213 (1): 15–22. ISSN 0021-9525. doi:10.1083/jcb.201511041.
Посилання |
Сайт присвячений Редагуванню РНК (англ.)
DARNED (DAtabase of RNa EDiting in humans, mice and flies). База даних з редагування РНК у людини, мишей і мух(англ.)
Сайт з редагування A на I(англ.)
Сайт з редагування C на U(англ.)
| ||||||||||||||||||||||||||||||||||||||||||||||
| Ця стаття належить до вибраних статей української Вікіпедії. |


